Full text loading...
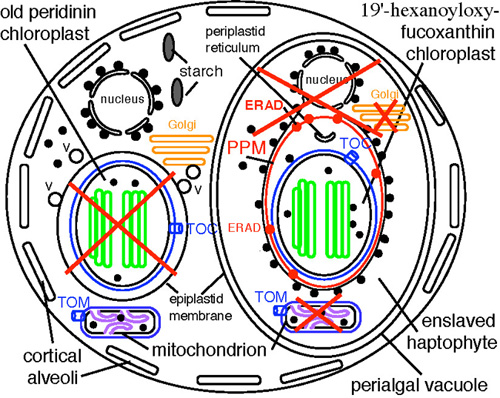
Symbiogenesis is the extremely rare, but permanent merger of two organisms from phylogenetically distant lineages into one radically more complex organism. Three examples are exceptionally important: intracellular enslavement by an early eukaryote of an α-proteobacterium by host protein insertion to make mitochondria; later conversion of a cyanobacterium into the first chloroplast, thereby forming kingdom Plantae; and secondary enslavement of a red alga to yield more complex membrane topology in the phagophototrophic kingdom Chromista. Two other cases involved independent acquisition of green-algal chloroplasts by ancestrally phagotrophic lineages, yielding chlorarachnean algae (phylum Cercozoa, within the chromist infrakingdom Rhizaria) and euglenophyte algae (phylum Euglenozoa, within the protozoan subkingdom Eozoa). Less radically, chloroplast replacement occurred within dinoflagellate Chromista by two symbiogeneses: Green-algal or haptophyte chloroplasts replaced ancestral peridinin-containing chloroplasts. These seven lineage mergers were all mediated by the evolution of novel modes of transmembrane protein import into the enslaved cell, allowing massive gene transfer from symbionts to host genomes.